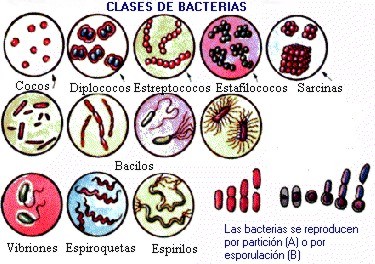
BACTERIAS
Las bacterias son microorganismos
unicelulares que presentan un tamaño de algunos micrómetros de largo (entre 0,5
y 5 um, por lo general) y diversas formas incluyendo esferas, barras y hélices.
Las bacterias son procariotas y, por lo tanto, no tienen núcleo ni orgánulos
internos. Generalmente poseen una pared celular compuesta de peptidoglucanos.
Muchas bacterias disponen de flagelos o de otros sistemas de desplazamiento y
son móviles.
Aunque el término bacteria incluía
tradicionalmente a todos los procariotas, actualmente la taxonomía y la
nomenclatura científica los divide en dos grupos. Estos dominios evolutivos se
denominan Bacteria y Archaea. La división se justifica en las grandes diferencias
que presentan ambos grupos a nivel bioquímico y en aspectos estructurales.
Los seres vivos se dividen
actualmente en tres dominios: bacterias (Bacteria), arqueas (Archaea) y
eucariontes (Eukarya). En los dominios Archaea y Bacteria se incluyen los
organismos procariotas, esto es, aquellos cuyas células no tienen un núcleo
celular diferenciado, mientras que en el dominio Eukarya se incluyen las formas
de vida más conocidas y complejas (protistas, animales, hongos y plantas).
Las bacterias también han estado implicadas en la segunda gran
divergencia evolutiva, la que separó Archaea de Eukarya. Se considera que las mitocondrias
de los eucariontes proceden de la endosimbiosis de una proteobacteria alfa. En
este caso, el antepasado de los eucariontes, que posiblemente estaba
relacionado con las arqueas (el organismo Neomura), ingirió una proteobacteria
que, al escapar a la digestión, se desarrolló en el citoplasma y dio lugar a
las mitocondrias. Estas se pueden encontrar en todos los eucariontes, aunque a
veces en formas muy reducidas, como en los protistas amitocondriales. Después,
e independientemente, una segunda endosimbiosis por parte de algún eucarionte
mitocondrial con una cianobacteria condujo a la formación de los cloroplastos
de algas y plantas. Se conocen incluso algunos grupos de algas que se han originado
claramente de acontecimientos posteriores de endosimbiosis por parte de eucariotas
heterótrofos que, tras ingerir algas eucariotas, se convirtieron en plastos de segunda
generación.
Morfología de las bacterias
Las bacterias presentan una amplia
variedad de tamaños y formas. La mayoría presentan un tamaño diez veces menor
que el de las células eucariotas, es decir, entre 0,5 y 5μm. Sin embargo,
algunas especies como Thiomargarita namibiensis y Epulopiscium fishelsoni llegan
a alcanzar los 0,5 mm, lo cual las hace visibles al ojo desnudo. En el otro
extremo se encuentran bacterias más pequeñas conocidas, entre las que cabe destacar
las pertenecientes al género Mycoplasma, las cuales llegan a medir solo 0,3 µm,
es decir, tan pequeñas como los virus más grandes. La forma de las bacterias es
muy variada y, a menudo, una misma especie adopta distintos tipos morfológicos,
lo 24 que se conoce como polimorfismo. De todas formas, podemos distinguir tres
tipos fundamentales de bacterias:
● Coco (del griego kókkos, grano): de
forma esférica.
○ Diplococo: cocos en grupos de dos.
○ Tetracoco: cocos en grupos de
cuatro.
○ Estreptococo: cocos en cadenas.
○ Estafilococo: cocos en agrupaciones
irregulares o en racimo.
● Bacilo (del latín baculus,
varilla): en forma de bastoncillo.
● Formas helicoidales:
○ Vibrio: ligeramente curvados y en
forma de coma, judía o cacahuete.
○ Espirilo: en forma helicoidal
rígida o en forma de tirabuzón.
○ Espiroqueta: en forma de tirabuzón
(helicoidal flexible).
Algunas especies presentan incluso
formas tetraédricas o cúbicas. Esta amplia variedad de formas es determinada en
última instancia por la composición de la pared celular y el citoesqueleto,
siendo de vital importancia, ya que puede influir en la capacidad de la
bacteria para adquirir nutrientes, unirse a superficies o moverse en presencia de
estímulos. A continuación se citan diferentes especies con diversos patrones de
asociación:
● Neisseria gonorrhoeae en forma
diploide (por pares).
● Streptococcus en forma de cadenas.
● Staphylococcus en forma de racimos.
● Actinobacteria en forma de
filamentos. Dichos filamentos suelen rodearse de una vaina que contiene
multitud de células individuales, pudiendo llegar a ramificarse, como el género
Nocardia, adquiriendo así el aspecto del micelio de un hongo.
Las bacterias presentan la capacidad
de anclarse a determinadas superficies y formar un agregado celular en forma de
capa denominado biopelícula o biofilme, los cuales pueden tener un grosor que
va desde unos pocos micrómetros hasta medio metro. Estas biopelículas pueden
congregar diversas especies bacterianas, además de protistas 24 y arqueas, y se
caracterizan por formar un conglomerado de células y componentes
extracelulares, alcanzando así un nivel mayor de organización o estructura
secundaria denominada microcolonia, a través de la cual existen multitud de
canales que facilitan la difusión de nutrientes. En ambientes naturales tales
como el suelo o la superficie de las plantas, la mayor parte de las bacterias
se encuentran ancladas a las superficies en forma de biopelículas. Dichas
biopelículas deben ser tenidas en cuenta en las infecciones bacterianas
crónicas y en los implantes médicos, ya que las bacterias que forman estas
estructuras son mucho más difíciles de erradicar que las bacterias individuales.
Por último, cabe destacar un tipo de morfología más compleja aún, observable en
algunos microorganismos del grupo de las mixobacterias. Cuando estas bacterias
se encuentran en un medio escaso en aminoácidos son capaces de detectar a las
células de alrededor, en un proceso conocido como quorum sensing, en el cual
todas las células migran hacia las demás y se agregan, dando lugar a cuerpos
fructíferos que pueden alcanzar los 0,5 mm de longitud y contener unas 100.000
células. Una vez formada dicha estructura las bacterias son capaces de llevar a
cabo diferentes funciones, es decir, se diferencian, alcanzando así un cierto
nivel de organización pluricelular. Por ejemplo, entre una y diez células
migran a la parte superior del cuerpo fructífero y, una vez allí, se
diferencian para dar lugar a un tipo de células latentes denominadas
mixosporas, las cuales son más resistentes a la desecación y, en general, a
condiciones ambientales adversas.
Las bacterias son organismos
relativamente sencillos. Sus dimensiones son muy reducidas, unos 2 μm de ancho
por 7-8 μm de longitud en la forma cilíndrica (bacilo) de tamaño medio; aunque
son muy frecuentes las especies de 0,5-1,5 μm. Carecen de un núcleo delimitado
por una membrana aunque presentan un nucleoide, una estructura elemental que
contiene una gran molécula circular de ADN. El citoplasma carece de orgánulos
delimitados por membranas y de las formaciones protoplasmáticas propias de las
células eucariotas. En el citoplasma se pueden apreciar plásmidos, pequeñas
moléculas circulares de ADN que coexisten con el nucleoide, contienen genes y
son comúnmente usados por las bacterias en la conjugación.
El 24 citoplasma también contiene
vacuolas (gránulos que contienen sustancias de reserva) y ribosomas (utilizados
en la síntesis de proteínas). Una membrana citoplasmática compuesta de lípidos
rodea el citoplasma y, al igual que las células de las plantas, la mayoría
posee una pared celular, que en este caso está compuesta por peptidoglicano
(mureína). Algunas bacterias, además, presentan una segunda membrana lipídica
(membrana externa) rodeando a la pared celular. El espacio comprendido entre la
membrana citoplasmática y la pared celular (o la membrana externa si esta
existe) se denomina espacio periplásmico. Algunas bacterias presentan una
cápsula y otras son capaces de evolucionar a endosporas, estadios latentes
capaces de resistir condiciones extremas. Entre las formaciones exteriores
propias de la célula bacteriana destacan los flagelos y los pili.
La membrana citoplasmática de las
bacterias es similar a la de plantas y animales, si bien generalmente no
presenta colesterol. Tiene una estructura similar a la de plantas y animales.
Es una bicapa lipídica compuesta fundamentalmente de fosfolípidos en la que se
insertan moléculas de proteínas. En las bacterias realiza numerosas funciones
entre las que se incluyen las de barrera osmótica, transporte, biosíntesis,
transducción de energía, centro de replicación de ADN y punto de anclaje para
los flagelos. A diferencia de las membranas eucarióticas, generalmente no
contiene esteroles (son excepciones micoplasmas y algunas proteobacterias),
aunque puede contener componentes similares denominados hopanoides. Muchas
importantes reacciones bioquímicas que tienen lugar en las células se producen
por la existencia de gradientes de concentración a ambos lados de una membrana.
Este gradiente crea una diferencia potencial análoga a la de una batería
eléctrica y permite a la célula, por ejemplo, el transporte de electrones y la
obtención de energía. La ausencia de membranas internas en las bacterias
significa que estas reacciones tienen que producirse a través de la propia
membrana citoplasmática, entre el citoplasma y el espacio periplásmico. Puesto
que las bacterias son procariotas no tienen orgánulos citoplasmáticos
delimitados por membranas y por ello presentan pocas estructuras
intracelulares. Carecen de núcleo celular, mitocondrias, cloroplastos y de los
otros orgánulos presentes en las células eucariotas, tales como el aparato de
Golgi y el retículo endoplasmático. Como excepción, algunas bacterias contienen
estructuras intracelulares rodeadas por membranas que pueden considerarse
primitivos orgánulos. Un ejemplo son los “tilacoides” de las cianobacterias.
Las bacterias contienen ribosomas para la síntesis de proteínas, pero éstos son
diferentes a los de eucariotas y arqueas. La estructura de los ribosomas de
arqueas y bacterias es 24 similar, pues ambos son de tipo 70S mientras que los
ribosomas eucariotas son de tipo 80S. Sin embargo, la mayoría de las proteínas
ribosomiales, factores de traducción y ARNt arqueanos son más parecidos a los
eucarióticos que a los bacterianos. Muchas bacterias presentan gránulos
intracelulares para el almacenaje de sustancias, como por ejemplo glucógeno, polifosfatos,
azufre o polihidroxialcanoatos.
Estructuras extracelulares
Las bacterias disponen de una pared celular que rodea a su membrana
citoplasmática. Las paredes celulares bacterianas están hechas de
peptidoglucano (llamado antiguamente mureína). Esta sustancia está compuesta
por cadenas de polisacárido enlazadas por péptidos inusuales que contienen
aminoácidos. Estos aminoácidos no se encuentran en las proteínas, por lo que
protegen a la pared de la mayoría de las peptidasas. Las paredes celulares bacterianas
son distintas de las que tienen plantas y hongos, compuestas de celulosa y
quitina, respectivamente. Son también distintas a las paredes celulares de
Archaea, que no contienen peptidoglucanos. El antibiótico penicilina puede
matar a muchas bacterias inhibiendo un paso de la síntesis del peptidoglucano.
Existen dos diferentes tipos de pared
celular bacteriana denominadas Gram-positiva y 24 Gram-negativa,
respectivamente. Estos nombres provienen de la reacción de la pared celular a
la tinción de Gram, un método tradicionalmente empleado para la clasificación
de las especies bacterianas. Las bacterias Gram-positivas tienen una pared
celular gruesa que contiene numerosas capas de peptidoglicano en las que se
inserta ácido teicoico. En cambio, las bacterias Gram-negativas tienen una
pared relativamente fina, consistente en unas pocas capas de peptidoglicano,
rodeada por una segunda membrana lipídica (la membrana externa) que contiene
lipopolisacáridos y lipoproteínas. Los micoplasmas son una excepción, pues
carecen de pared celular. La mayoría de las bacterias tienen paredes celulares
Gram-negativas; solamente son Gram-positivas Firmicutes y Actinobacteria. Estos
dos grupos eran antiguamente conocidos como bacterias Gram-positivas de
contenido GC bajo y bacterias Gram-positivas de contenido GC alto,
respectivamente. Estas diferencias en la estructura de la pared celular dan
lugar a diferencias en la susceptibilidad antibiótica. Por ejemplo, la
vancomicina puede matar solamente a bacterias Gram-positivas y es ineficaz
contra patógenos Gramnegativos, tales como Haemophilus influenzae o Pseudomonas
aeruginosa. Dentro del filo Actinobacteria cabe hacer una mención especial al
género Mycobacterium, el cual, si bien se encuadra dentro de las Gram
positivas, no parece serlo desde el punto de vista empírico, ya que su pared no
retiene el tinte. Esto se debe a que presentan una pared celular poco común,
rica en ácidos micólicos, de carácter hidrófobo y ceroso y bastante gruesa, lo
que les confiere una gran resistencia.
Los flagelos son largos apéndices
filamentosos compuestos de proteínas y utilizados para el movimiento. Tienen un
diámetro aproximado de 20 nm y una longitud de hasta 20 μm. Los flagelos son
impulsados por la energía obtenida de la transferencia de iones. Esta
transferencia es impulsada por el gradiente electroquímico que existe entre
ambos lados de la membrana citoplasmática.
Endosporas
Ciertos géneros de bacterias
Gram-positivas, tales como Bacillus, Clostridium, Sporohalobacter, Anaerobacter
y Heliobacterium, pueden formar endosporas. Las endosporas son estructuras
durmientes altamente resistentes cuya función primaria es sobrevivir cuando las
condiciones ambientales son adversas. En casi todos los casos, las endosporas
no forman parte de un proceso reproductivo, aunque Anaerobacter puede formar
hasta siete endosporas a partir de una célula. Las endosporas tienen una base
central de citoplasma que contiene ADN y ribosomas, rodeada por una corteza y
protegida por una cubierta impermeable y rígida. Las endosporas no presentan un
metabolismo detectable y pueden sobrevivir a condiciones físicas y químicas
extremas, tales como altos niveles de luz ultravioleta, rayos gamma,
detergentes, desinfectantes, calor, presión y desecación. En este estado
durmiente, las bacterias pueden seguir viviendo durante millones de años, e
incluso pueden sobrevivir en la radiación y vacío del espacio exterior. Las
endosporas pueden también causar enfermedades. Por ejemplo, puede contraerse
carbunco por la inhalación de endosporas de Bacillus anthracis y tétanos por la
contaminación de las heridas con endosporas de Clostridium tetani.
Metabolismo
En contraste con los organismos superiores,
las bacterias exhiben una gran variedad de tipos metabólicos. La distribución
de estos tipos metabólicos dentro de un grupo de bacterias se ha utilizado
tradicionalmente para definir su taxonomía, pero estos rasgos no corresponden a
menudo con las clasificaciones genéticas modernas. El metabolismo bacteriano se
clasifica en base a tres criterios importantes: el origen del carbono, la
fuente de energía y los donadores de electrones. Un criterio adicional para
clasificar a los microorganismos que respiran es el receptor de electrones
usado en la respiración. Según la fuente de carbono, las bacterias se pueden
clasificar como:
● Heterótrofas, cuando usan compuestos
orgánicos.
● Autótrofas, cuando el carbono celular se
obtiene mediante la fijación del dióxido de carbono. Las bacterias autótrofas
típicas son las cianobacterias fotosintéticas, las bacterias verdes del azufre
y algunas bacterias púrpura. Pero hay también muchas otras especies
quimiolitotrofas, por ejemplo, las bacterias nitrificantes y oxidantes del
azufre. Según la fuente de energía, las bacterias pueden ser:
● Fototrofas, cuando emplean la luz a
través de la fotosíntesis.
● Quimiotrofas, cuando obtienen
energía a partir de sustancias químicas que son oxidadas principalmente a
expensas del oxígeno (respiración aerobia) o de otros receptores de electrones
alternativos (respiración anaerobia). Según los donadores de electrones, las
bacterias también se pueden clasificar como: 24
● Litotrofas, si utilizan como
donadores de electrones compuestos inorgánicos.
● Organotrofas, si utilizan como
donadores de electrones compuestos orgánicos. Los organismos quimiotrofos usan
donadores de electrones para la conservación de energía (durante la respiración
aerobia, anaerobia y la fermentación) y para las reacciones biosintéticas (por
ejemplo, para la fijación del dióxido de carbono), mientras que los organismos
fototrofos los utilizan únicamente con propósitos biosintéticos. Los organismos
que respiran usan compuestos químicos como fuente de energía, tomando
electrones del sustrato reducido y transfiriéndolos a un receptor terminal de
electrones en una reacción redox. Esta reacción desprende energía que se puede
utilizar para sintetizar ATP y así mantener activo el metabolismo.
Movimiento
Algunas bacterias son inmóviles y
otras limitan su movimiento a cambios de profundidad. Por ejemplo,
cianobacterias y bacterias verdes del azufre contienen vesículas de gas con las
que pueden controlar su flotabilidad y así conseguir un óptimo de luz y alimento.
Las bacterias móviles pueden desplazarse por deslizamiento, mediante
contracciones o más comúnmente usando flagelos. Algunas bacterias pueden
deslizarse por superficies sólidas segregando una sustancia viscosa, pero el
mecanismo que actúa como propulsor es todavía desconocido. En el movimiento
mediante contracciones, la bacteria usa su pilus de tipo IV como gancho de
ataque, primero lo extiende, anclándolo y después lo contrae con una fuerza
notable (>80 pN). El flagelo bacteriano es un largo apéndice filamentoso
helicoidal propulsado por un motor rotatorio (como una hélice) que puede girar
en los dos sentidos. El motor utiliza como energía un gradiente electroquímico
a través de la membrana. Los flagelos están compuestos por cerca de 20
proteínas, con aproximadamente otras 30 proteínas para su regulación y
coordinación. Hay que tener en cuenta que, dado el tamaño de la bacteria, el
agua les resulta muy viscosa y el mecanismo de propulsión debe ser muy potente
y eficiente. Los flagelos bacterianos se encuentran tanto en las bacterias
Gram-positivas como Gram-negativas y son completamente diferentes de los
eucarióticos y, aunque son superficialmente similares a los arqueanos, se
consideran no homólogos.
Las bacterias crecen hasta un tamaño
fijo y después se reproducen por fisión binaria una forma de reproducción
asexual.
Faces de las bacterias:
- Fase de adaptación.
- Fase exponencial.
- Fase estacional.